Towards virtual meristem
During the last few years a significant progress has been made in the understanding of meristem development. In a first phase, a focus has been put on the analysis of individual processes. Due to this fact, an integrated view which would connect different parts of the current state of art is currently not possible. The partial solution of this problem might be the systematic application of currently accessible data in a computer models. Modelling usually connects the domains, helps in classifying data and shows the weak points of our understanding.
The main goal of my work withing Sy-stem project is to develop a model of apical meristem which could be used to make in silico experiments. The main advantages of making virtual meristems are:
- possibility to verify our understanding of current state of art -- by introducing hypotheses to the model we may test their completeness (which means answering the question: does our current understanding let us fully explain observed phenomena?) and correctness (if we have a complete set of axioms and we introduced valid hypotheses, is the model behaviour similar to the one observed in real world),
- possibility to test new hypotheses -- by introducing new hypothesis we may observe the model behaviour and check if the hypothesis is valid in the model. If so, we can plan an experiment on real object to test the validity of hypothesis in this object.
- although testing and verifying of hypotheses is also possible in real experiments (without modelling), proceeding with in silico experiment is often faster and cheaper.
Another goal of my work within Sy-stem is to investigate the methodology of simulating cell tissue systems as Dynamic Systems with a Dynamic Structure (DS2). The developing tissue may be viewed as a dynamic system (which means that the system could be described by states, changing in the time) with a dynamic topology (also changing in the time). We call such structures DS2 systems. To simulate that class of systems special mechanisms should be used. An analysis of influence of these mechanisms on the simulation result could help in creating methodology of cell tissue systems.The main difficulty of creating a meristem model are:
- big amount of available data -- currently we have a lot of accessible data describing the meristem at different scales (eg. genes activity, molecules transport, geometry describing the whole meristem, etc.). During last period an important progress in understanding of meristem functioning has been made. However, it still lacks answers for many important questions (eg. when exactly the symmetry of young meristem is broken? why the phyllotactic patterns are formed?).
- multiple processes with feedback loops -- even for many processes which were partly understood the relation between cause and effect is not precisely defined (eg. does the growth imply the physiology or does the physiology imply the growth or is there a feedback from one to another?).
- modeling the complete physiology (including the link between biomechanics and biochemistry). The shape of the meristem is the result of mechanical properties of material and growth. The other components of physiology (like signalling, metabolism, transport, regulation) are described mainly by chemical processes. According to our current knowledge, finding the connection between these two domains (mechanics and chemistry) at big scale level is not extensively investigated and seems to be tough task. In addition, the mathematical tools, which could be used to describe the shape transformations are either difficult to use (eg. tensor mechanics) or inaccurate, giving big errors (eg. shape analysis based on solving problems from optimisation domain). The fact, that the chemical nature of meristem is hard to transfer from small to big scale is an additional problem.
Results
Here some preliminary results can be found:
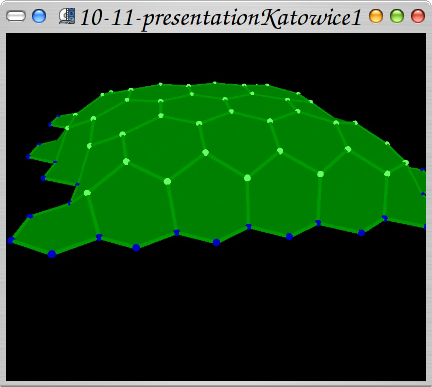
Model showing a L1 and inside representation. The model and all submodels are besed on WalledTissue structure, which was created as a part of my PhD thesis and as a support for VirtualPlants team.
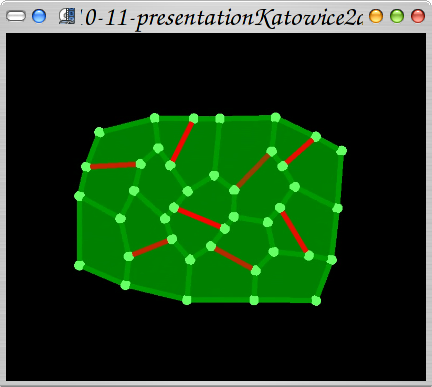
Poligonal mesh division algorithm with parameters tuned to reflect biological phenomena. It is used to simulate cell divisions in meristem tissues.
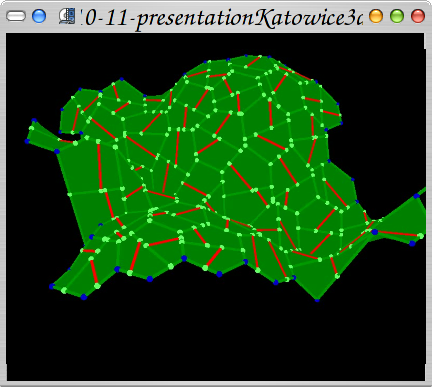
Same algorithm used to create dense polygonal mesh of cells.
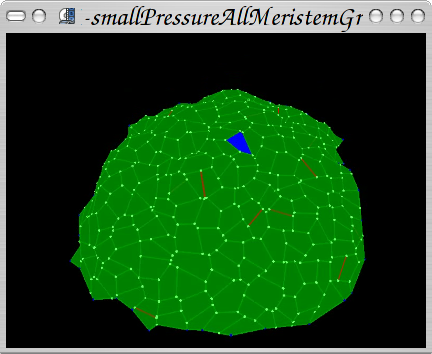
Growth algorithm based on manipulating wall length applied to real meristem digitalized by Pierre Barbier de Reuille's ;] protocol of 4D meristem reconstruction.
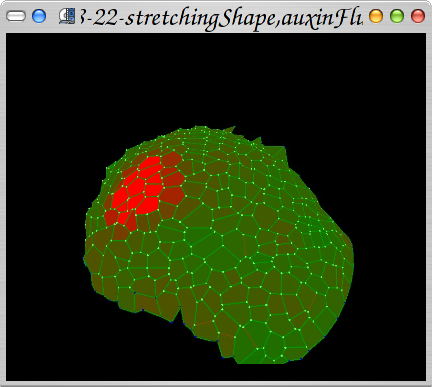
Simple application of physiology process in the meristeme tissue: factor X diffusion.
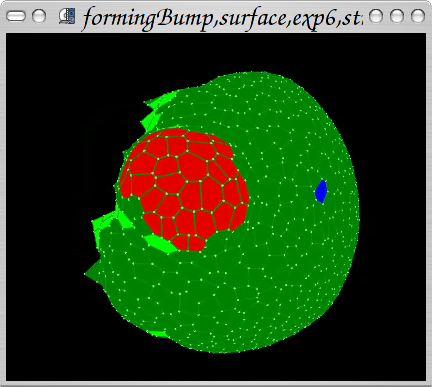
First steps in coupling mechanics with physiology: reproduction of PIN1 mutant behaviour with hypotetic 'stress' gene expression. Auxin: red. Light green: stress gene expressed.
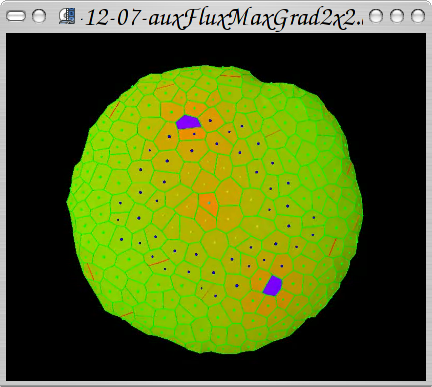
First steps in phyllotaxis based on local rules: implementation of MaxGradient pump orientation hypothesis.
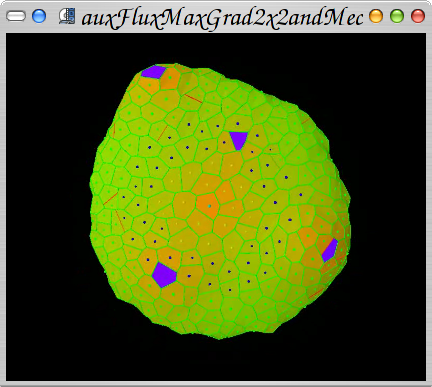
First steps in coupling mechanics with phyllotactic patterns.
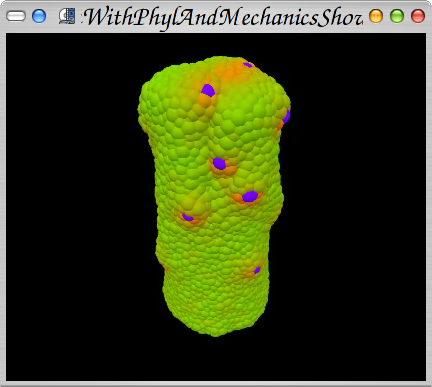
Results of longth growth with coupled mechanics and auxin transport showing the problem of initial cell placement.
Results of longth growth with coupled mechanics and auxin transport showing the problem of initial cell placement.
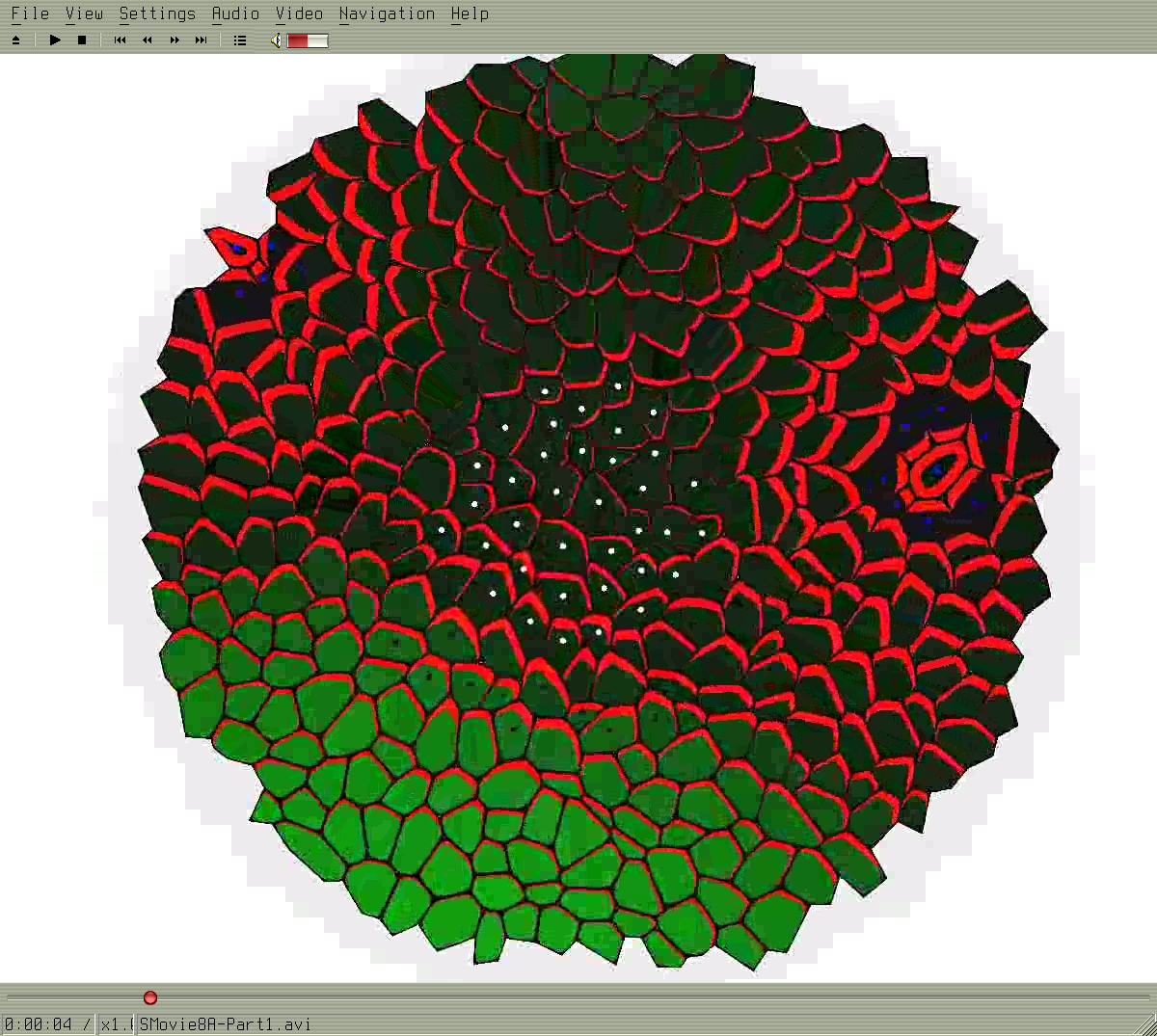
SMovie8A-Part1.avi
Formation of spiral phyllotaxis pattern. The inhibitory field concept is realized using the auxin concentration gradient. The auxin is distributed using an active transport. The active transport molecules (PINs) are distributed with concordance to the canalization hypothesis.
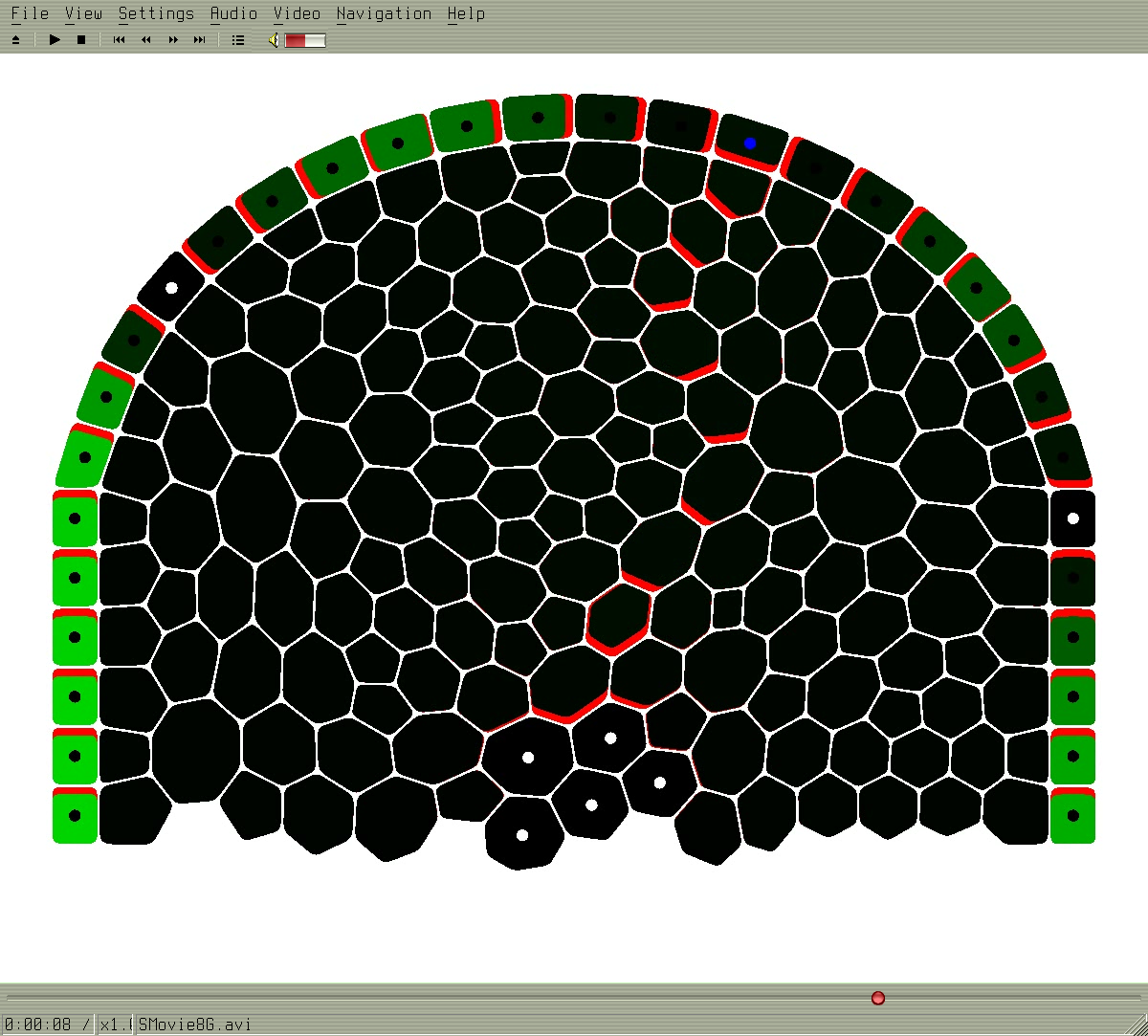
SMovie8G.avi
Formation of vascular strain below primordia. Another canalization example. L1 works also with canalization: the difference lay in the assumption of auxin creation and different Phi function.

SMovie9A.avi
Root system with the canalization hypohesis.